Apariția noului SARS-CoV-2 de coronavirus uman la Wuhan, China, a provocat o epidemie mondială de boli respiratorii (COVID-19). În prezent, lipsesc vaccinurile și terapeuții orientate pentru tratamentul acestei boli. Aici raportăm un anticorp monoclonal uman care neutralizează SARS-CoV-2 (și SARS-CoV) în cultura celulară. Acest anticorp neutralizant încrucișat vizează un epitop comun asupra acestor viruși și poate oferi potențial pentru prevenirea și tratarea COVID-19.
Introducere
Sindromul respirator acut sever coronavirus 2 (SARS-CoV-2) este agentul etiologic al bolii 19 (COVID-19) indusă de coronavirus care a apărut în China la sfârșitul lui 2019 și a cauzat o pandemie 1 . La 19 aprilie 2020, 2.241.778 de cazuri au fost raportate la nivel mondial, dintre care 152.551 (6,8%) au cedat infecției 2 . SARS-CoV-2 aparține subgenului Sarbecovirus (genul Betacoronavirus , familia Coronaviridae ) 3împreună cu SARS-CoV care a apărut în 2002 provocând ~ 8000 de infecții cu o letalitate de 10%. Ambele virusuri au traversat barierele speciilor dintr-un rezervor de animale și pot cauza o boală respiratorie care poate pune viața la om. În prezent, nu există disponibile terapeutice vizate pentru COVID-19. Anticorpii monoclonali care vizează site-urile vulnerabile pe proteinele de suprafață virale sunt recunoscuți din ce în ce mai mult ca o clasă promițătoare de medicamente împotriva bolilor infecțioase și au demonstrat eficacitate terapeutică pentru un număr de viruși 4 , 5 .
Anticorpi de neutralizare a coronavirusului vizează în primul rând glicoproteinele spice (S) trimeric pe suprafața virală care mediază intrarea în celulele gazdă. Proteina S are două subunități funcționale care mediază atașarea celulelor (subunitatea S1, existentă din patru domenii de bază S1 A până la S1 D ) și fuziunea membranei virale și celulare (subunitatea S2). Anticorpi de neutralizare a potențialului vizează adesea locul de interacțiune cu receptorii în S1, dezactivând interacțiunile cu receptorii 6 , 7 , 8 , 9 , 10 , 11. Proteinele spike ale SARS-CoV-2 (SARS2-S; 1273 reziduuri, tulpină Wuhan-Hu-1) și SARS-CoV (SARS-S, 1255 reziduuri, tulpină Urbani) sunt 77,5% identice prin secvența de aminoacizi primari, sunt foarte similar 12 din punct de vedere structural , 13 , 14 , 15 și se leagă în mod obișnuit de proteina enzimă 2 (ACE2) protejată de angiotensină ca receptor gazdă 1 , 16 prin domeniul lor S1 B. Se știe că interacțiunea dintre receptori declansează modificări conformaționale ireversibile ale proteinelor spike coronavirus care permit fuziunea membranei 17 .
Rezultate
Identificarea anticorpilor reactivi SARS-CoV-2
Pentru a identifica anticorpii de neutralizare a SARS-CoV-2, a fost evaluată reactivitatea ELISA- (încrucișată) a supernatanților care conțin anticorpi dintr-o colecție de 51 hibridoma SARS-S derivată din șoareci transgenici H2L2 transgenici imunizați care codifică imunoglobuline himerice cu variabilă umană grea și lanțuri ușoare și regiuni constante de origine șobolan (Tabelul suplimentar 1 ). Patru din 51 de supernatante de hibridom SARS-S au afișat reactivitatea ELISA-încrucișare cu subunitatea SARS2-S1 (reziduurile S 1-681; Tabelul suplimentar 1), dintre care unul (47D11) a prezentat o activitate de neutralizare încrucișată a infecției VSV pseudotipate SARS-S și SARS2-S. Anticorpul himeric 47D11 H2L2 a fost reformatat într-o imunoglobulină complet umană, prin donarea regiunilor variabile umane grele și cu lanț ușor într-o coloană vertebrală izotip IgG1 uman. 47D11 uman exprimat recombinant a fost utilizat pentru caracterizarea ulterioară.
Proprietăți antivirale și biochimice ale mAb 47D11 uman
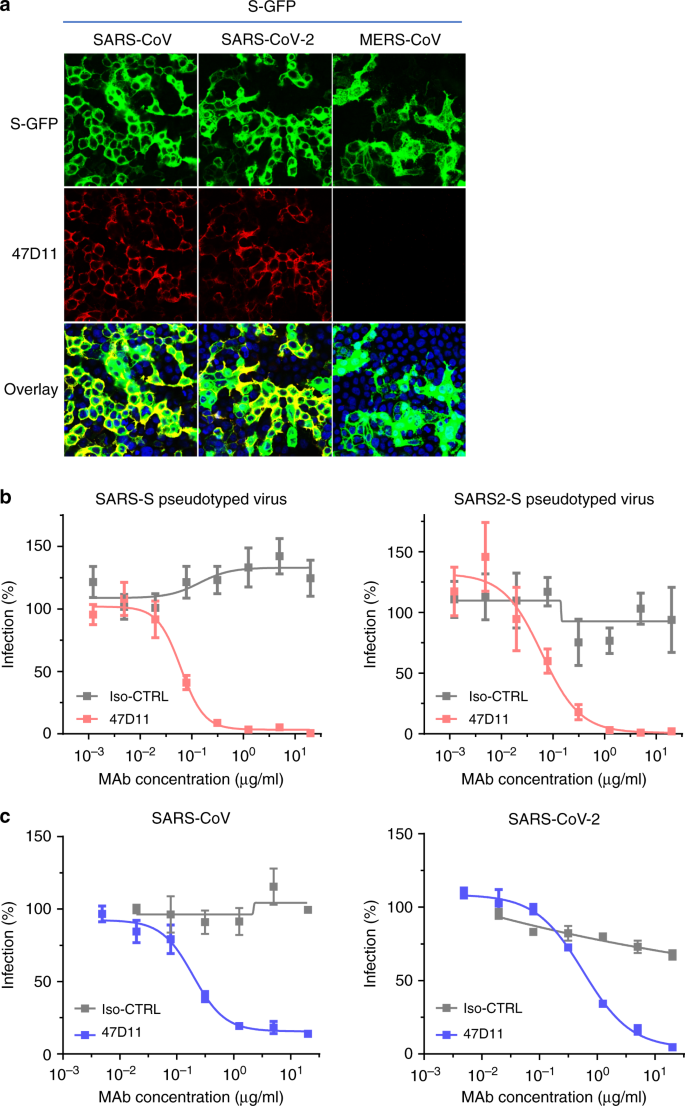
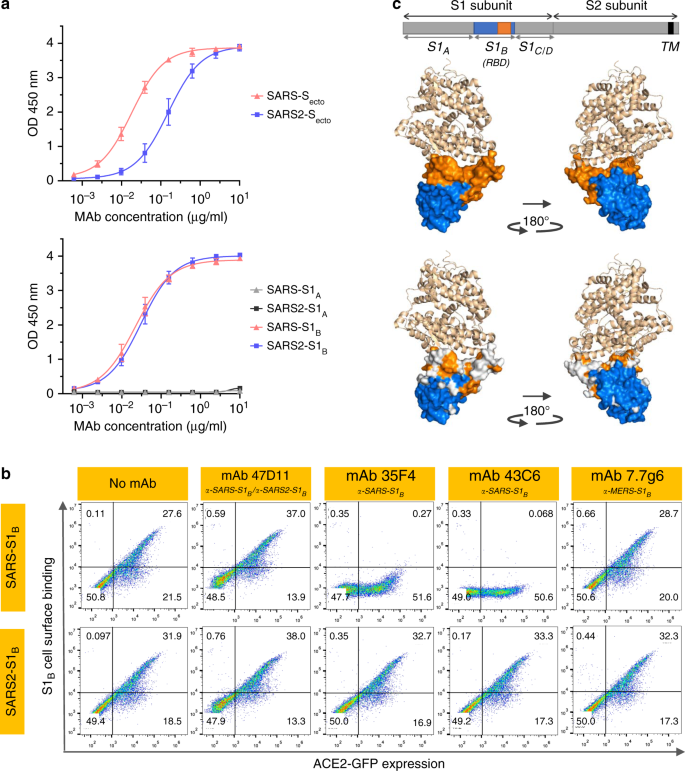
Anticorpul uman 47D11 se leagă la celule care exprimă proteinele spike de lungime întreagă ale SARS-CoV și SARS-CoV-2 (Fig. 1a ). Anticorpul 47D11 a fost descoperit infecția celulelor inhiba puternic VeroE6 cu SARS-S și SARS2-S pseudotyped VSV cu IC 50 de valori de 0,061 și 0,061 pg / ml (Fig. 1b ), respectiv. Infecție autentică a celulelor VeroE6 cu SARS-CoV și SARS-CoV-2 a fost neutralizat cu IC 50 de valori de 0,19 și 0,57 pg / ml (Fig. 1c ). Utilizarea ELISA 47D11 s-a dovedit că vizează domeniul de legare la receptorii S1 B (RBD) al SARS-S și SARS2-S. 47D11 a legat S1 B de ambele virusuri cu afinități similare, așa cum se arată în concentrația efectivă jumătate maximă bazată pe ELISA (CE50 ) valori (0,02 și respectiv 0,03 μg / ml; Fig. 2a ). Afinitatea de legare bazată pe ELISA, 47D11, pentru ectodomina de vârf (S ecto ) a SARS-CoV a fost mai mare comparativ cu cea a SARS-CoV-2 ( valori EC 50 : 0,018 și, respectiv, 0,15 μg / ml), în ciuda acoperirii antigenului echimolar ( Fig. 1 ). Congruent cu reactivitățile ELISA, măsurarea cineticii de legare a 47D11 prin interferometrie cu biocaparat a arătat că 47D11 leagă SARC-S ecto cu afinitate mai mare (constanta de disociere a echilibrului [ K D ]: 0,745 nM) în raport cu ecto SARS2-S ( K D 10,8 nM) ), în timp ce afinitatea pentru SARS-S1 Bși SARS2-S1 B se găseau într-un interval similar (16,1 și, respectiv, 9,6 nM, Fig. 2 suplimentară ). Această diferență poate provoca din diferențele de accesibilitate a epitopului în SARS-S față de SARS2-S, întrucât domeniul B poate adopta o conformare închisă și deschisă în omotrimerul 12 , 13 al punctului de preferință . Remarcabil, legarea 47D11 la SARS-S1 B și SARS2-S1 B nu a concurat cu legarea S1 B la receptorul ACE2 exprimat la suprafața celulei așa cum se arată prin citometria în flux (Fig. 2b ; Fig. 3 suplimentară ) și nici cu Secto și S1 Bcare se leagă de ACE2 solubilă în test pe bază de fază solidă (Fig. 4 suplimentară ), în timp ce doi anticorpi specifici SARS-S1 35F4 și 43C6 care neutralizează SARS-S (dar nu SARS2-S) infecție VSV pseudotipată (Fig. 5 suplimentară ) blochează legarea SARS-S ecto și SARS-S1 B la ACE2. Folosind un test de fuziune celulă declanșată prin trypsină, s-a demonstrat că 47D11 afectează formarea de sincitie SARS-S și SARS2-S (Fig. Suplimentară Fig. 6). Datele noastre arată că 47D11 neutralizează SARS-CoV și SARS-CoV-2 printr-un mecanism încă necunoscut care este diferit de interferența care leagă receptorul. Au fost raportate mecanisme alternative de neutralizare a coronavirusului prin anticorpi care vizează RBD, incluzând inactivarea vârfului prin destabilizarea indusă de anticorp a structurii sale de prefuzie 17 , care poate fi aplicată și pentru 47D11.
Fig. 1: 47D11 neutralizează SARS-CoV și SARS-CoV-2.
o legare de 47D11 la celulele HEK-293T care exprimă proteinele spike marcate cu GFP ale SARS-CoV și SARS-CoV-2 detectate prin testul de imunofluorescență. MAb 7.7G6 uman care vizează domeniul de vârf MERS-CoV S1 B a fost luat ca un control negativ, nucleele de celule din imaginile suprapuse sunt vizualizate cu DAPI. bNeutralizarea mediată de anticorpi a infecției particulelor VSV care codifică luciferaza pseudotipate cu proteine spike ale SARS-CoV și SARS-CoV-2. Particule pseudotipate de VSV pre-incubate cu anticorpi la concentrații indicate (a se vedea metodele) au fost utilizate pentru a infecta celulele VeroE6 și activitățile luciferazei în lizații celulari au fost determinate la 24 h post transducție pentru a calcula infecția (%) în raport cu controalele ne-tratate cu anticorp. Este prezentată media ± SD din cel puțin trei experimente independente cu triplicate tehnice. Iso-CTRL: un anticorp monoclonal 11 anti-Strep-tag uman a fost utilizat ca control al izotipului anticorpului. cNeutralizarea mediată de anticorpi a infecției cu SARS-CoV și SARS-CoV-2 pe celulele VeroE6. Experimentul a fost efectuat cu probe triplicate, se arată media ± SD. Datele sursă sunt furnizate ca fișier Date sursă.
Fig. 2: neutralizarea 47D11 mAb leagă SARS1-S și SARS2-S RBD fără a elimina interacțiunea dintre receptori.
un test ELISA de legare la curbele de 47D11 la S ecto (panou superior) sau S1 A si S1 B (RBD: domeniu de legare a receptorului) (panoul inferior) CoV-S și SARS2-S filmate la concentrații echimolare. Este prezentată media ± SD din două experimente independente cu duplicate tehnice. b Interferența anticorpilor cu legarea S-S1 B a SARS-CoV și SARS-CoV-2 la suprafața celulară ACE2-GFP analizată prin citometrie de flux. Înainte de legarea celulelor, S1 B a fost amestecat cu mAb (mAbs 47D11, 35F4, 43C6, 7.7G6, în format H2L2) cu specificitate indicată într-un raport molar mAb: S1 B de 8: 1 (a se vedea Fig. 3 suplimentară pentru o extensivă analiză folosind mAb diferit: S1Raportul molar B ). Celulele sunt analizate pentru expresia (ACE2-) GFP ( axa x ) și legarea S1 B ( axa y ). Procentele de celule care au obținut negativ, pozitiv unic sau dublu pozitiv sunt afișate în fiecare cadran. Experimentul a fost făcut de două ori, este prezentat un experiment reprezentativ. c Divergența reziduurilor de suprafață în S1 B din SARS-CoV și SARS-CoV-2. Panoul superior: Structura proteinei spice S1 B RBD SARS-CoV în complex cu receptorul ACE2 uman (PDB: 2AJF) 24 . ACE2 (culoarea grâului) este vizualizată în prezentarea panglicii. S1 Bdomeniul principal (albastru) și subdomeniul (portocaliu) sunt afișate în prezentarea suprafeței folosind PyMOL și sunt vizualizate cu aceleași culori în diagrama liniară a proteinei spike de mai sus, cu pozițiile subunităților S1 și S2, ectodomina S (S ecto ), domeniile S1 S1 A-D și domeniul transmembran (TM) indicate. Panou inferior: similar cu panoul de mai sus, cu reziduuri de suprafață pe S1 B din SARS-CoV, care diferă cu SARS-CoV-2, colorat în alb. Datele sursă sunt furnizate ca fișier Date sursă.
47D11 obiective un epitop conservat în SARS2-S-S1 B domeniu
SARB2-S1 B RBD (reziduurile 338–506) constă dintr-un domeniu de bază și un subdomeniu de legare a receptorilor (reziduuri 438-498) care se extind din structura de domeniu principal de betasheet antiparalel care angajează direct receptorul. Comparativ cu domeniul principal S1 B , identitatea secvenței proteice a receptorului S1 B care interacționează subdomeniul SARS-S și SARS2-S este substanțial mai mică (46,7% față de 86,3%; Fig. 7 și Fig. 2c ). Anticorpi neutralizanți potențiali vizează adesea acest subdomeniu de legare a receptorului. Cu toate acestea, datorită variațiilor obișnuite în acest subdomeniu, acești anticorpi sunt adesea virus-specific și se leagă și neutralizează slab virusurile 18 , 19. Caracterul reactiv încrucișat al 47D11 indică faptul că anticorpul este mai probabil să țintească structura de bază conservată a RBD S1 B. Interesant, anticorpul neutralizant SARS-CoV CR3022 care vizează și domeniul de bază al S1 B a fost găsit recent că se leagă încrucișat de SARS-CoV-2, deși nu a fost raportată capacitatea sa de a neutraliza încrucișat infecția SARS-CoV-2 18 , 20 . S1 Blegarea prin 47D11 mai departe de interfața receptor-legare explică incapacitatea sa de a compromite interacțiunea spike-receptor și deschide posibilități pentru tratamente combinate cu anticorpi neutralizanți necompetenți, puternici care vizează subdomeniul de legare la receptor. Combinațiile de anticorpi care vizează epitopii care nu se suprapun pot acționa sinergic ducând la o doză mai mică și pot diminua riscul de scăpare imună 20 .
În concluzie, acesta este primul raport al unui anticorp (uman) monoclonal care neutralizează SARS-CoV-2. 47D11 leagă un epitop conservat pe RBD-ul spike explicând capacitatea sa de a neutraliza încrucișat SARS-CoV și SARS-CoV-2, folosind un mecanism care este independent de inhibarea legării receptorilor. Acest anticorp va fi util pentru dezvoltarea testelor de detecție a antigenului și teste serologice care vizează SARS-CoV-2. Anticorpi neutralizanți pot modifica cursul infecției în gazda infectată care susține eliminarea virusului sau pot proteja o gazdă neinfectată care este expusă virusului 4 . Prin urmare, acest anticorp - fie singur, fie în combinație - oferă potențialul de a preveni și / sau trata COVID-19 și, de asemenea, și alte boli emergente viitoare la om cauzate de viruși din subgenul Sarbecovirus .
metode
Exprimarea și purificarea proteinelor spike de coronavirus
Ectodominanțele cu vârf de coronavirus (S ecto ) ale SARS-CoV-2 (reziduurile 1–1213; tulpina Wuhan-Hu-1; GenBank: QHD43416.1) și HCoV-OC43 (reziduuri 15–1263; tulpina Paris; UniProtKB: Q696P8 ) au fost exprimată tranzitoriu în celulele HEK-293T cu un motiv de trimerizare C-terminal și Strep-tag folosind plasmida de expresie pCAGGS. În mod similar, vectorii de expresie pCAGGS care codifică S1 sau subdomeniile sale de SARS-CoV (S1, reziduuri 1–676; S1 A , reziduuri 1-302; S1 B , reziduuri, 325–533) și SARS-CoV-2 (S1, reziduuri 1–682; S1 A , reziduuri 1–294; S1 B , reziduuri 329–538) Etichetatele terminale C cu domeniul Fc al IgG uman sau al mouse-ului sau cu tag-ul strep s-au generat așa cum este descris înainte de 21. Ectodomain spike Coronavirus din MERS-CoV (reziduurile 19–1262; tulpina EMC; GenBank: YP_009047204.1) și SARS-CoV (reziduuri 15–1182; tulpina Urbani; GeneBank: AY278741.1 ) fuzionate cu un motiv de trimerizare C-terminal, un site de clivaj de trombină și o etichetă de purificare a strep-tag-urilor au fost donate în cadru în pMT \ Bip \ V5 \ Vectorul său de expresie. Locul de clivaj al furinei la joncțiunea S1 / S2 a fost mutat pentru a preveni clivajul de furin în această poziție. Ectodomenii spike au fost produși stabil în linia celulară Drosophila S2, așa cum s-a descris anterior 22. Proteinele recombinante au fost purificate prin afinitate din supernatantul culturii prin purificarea de mărgele proteine A-sefarază (GE Healthcare, Catalog nr. 17-0780-01) sau perle de streptactin (IBA, Catalog nr. 2-1201-010). Puritatea și integritatea tuturor proteinelor recombinate purificate au fost verificate prin SDS-PAGE colorată de coomassie.
Generarea de H2L2 mAbs
Șoarecii H2L2 au fost imunizați secvențial la intervale de 2 săptămâni cu Secto purificatde diferite COV-uri în următoarea ordine: HCoV-OC43, SARS-CoV, MERS-CoV, HCoV-OC43, SARS-CoV și MERS-CoV. S-au injectat antigeni la 20-25 μg / mouse folosind Stimune Adjuvant (Prionics) proaspăt preparat conform instrucțiunilor producătorului pentru prima injecție, în timp ce stimularea s-a făcut cu ajutorul adjuvantului Ribi (Sigma). Injecțiile s-au făcut subcutanat în zona inghinală stângă și dreaptă fiecare (50 µl) și 100 µl intraperitoneal. La patru zile de la ultima injecție, se recoltează splina și ganglionii limfatici, iar hibridoamele sunt făcute prin metoda standard folosind linia celulară de mielom SP 2/0 (ATCC # CRL-1581) ca partener de fuziune. Hibridoamele au fost analizate în ELISA specifică antigenului și în cele selectate pentru dezvoltare ulterioară, subclonate și produse la scară mică (100 ml de mediu). În acest scop, hibridoamele sunt cultivate într-un mediu fără ser și proteine pentru cultivarea hibridomului (PFHM-II (1 ×), Gibco) cu adăugarea de aminoacizi neesențiali 100 × NEAA, Biowhittaker Lonza, Catalogul BE13-114E). Anticorpii H2L2 au fost purificați din supernatanții culturii de hibridom folosind cromatografia de afinitate Protein-G (Merck KGaA, Catalog nr. 16-266). Anticorpii purificați au fost depozitați la 4 ° C până la utilizare. Studiile la animale au fost realizate în baza permisului animal AVD101002016512, aprobat de CCD (comitetul central pentru experimente pe animale).
Producția de anticorp monoclonal uman 47D11
Pentru producția de mAb uman recombinant, codul ADNc care codifică regiunile variabile 47D11 H2L2 mAb ale lanțurilor grele și ușoare au fost donate în plasmide de expresie care conțin lanțuri grele IgG1 umane și, respectiv, kappa Ig Chapa, regiuni constante (InvivoGen). Ambele plasmide conțin secvența de semnal interleukin-2 pentru a permite secreția eficientă de anticorpi recombinanti. 47D11 mAb uman recombinant și controlul izotipului descris anterior (anti-strep-tag mAb) sau 7.7G6 mAb au fost produse în celulele HEK-293T în urma transfecției cu perechi de plasmide IgG1 cu catena grea și ușoară în conformitate cu protocoalele de la InvivoGen. Anticorpii umani au fost purificați din supernatanții culturii celulare folosind cromatografia de afinitate Protein-A. Anticorpii purificați au fost depozitați la 4 ° C până la utilizare.
Microscopie de imunofluorescență
Legarea anticorpilor la proteinele spike ale suprafeței celulare a SARS-CoV, SARS-CoV-2 și MERS-CoV a fost măsurată prin microscopie de imunofluorescență. Celulele HEK-293T (ATCC # CRL-3216), însămânțate pe lamelele de sticlă, au fost transfectate cu plasmide care codifică SARS-S, SARS2-S sau MERS-S - C-fuzibil C-terminal la proteina fluorescență verde (GFP) folosind Lipofectamina 2000 (Invitrogen , Catalog nr. 11668019). După două zile de la transfecție, celulele au fost fixate prin incubare cu 2% paraformaldehidă în soluție salină tamponată cu fosfat (PBS) timp de 20 min la temperatura camerei și colorate pentru nuclee cu 4,6-diamidino-2-fenilindol (Sigma, Catalog nr. D9542). Ulterior, celulele au fost incubate cu mAbs la o concentrație de 10 ug / ml timp de 1 oră la temperatura camerei, urmată de incubare cu anticorpi anti-IgG conjugati Alexa Fluor 594 conjugați de capră anti-IgG umană (Invitrogen, Thermo Fisher Scientific, Catalog nr. A-11014) timp de 45 min la temperatura camerei. Imaginile fluorescente au fost înregistrate folosind un microscop confocal Leica SpeII.
Test de inhibiție de legare a receptorului bazat pe citometrie
Interferența anticorpului de legătură S1 B la receptorul ACE2 uman pe suprafața celulelor a fost măsurată prin citometrie de flux. Celulele HEK-293T au fost însămânțate la o densitate de 2,5 × 10 5 celule per ml într-un balon T75. După atingerea confluenței 70-80%, celulele au fost transfectate cu o plasmidă de expresie care codifică ACE2 umană - fuzibilă C-terminal la GFP folosind Lipofectamina 2000 (Invitrogen). Două zile după transfecție, celulele au fost disociate prin soluție de disociere a celulelor (Sigma-aldrich, Merck KGaA; Catalog # C5914). În total, 2,5 pg / ml de FC uman marcate SARS-S1 B și SARS2-S1 B au fost pre-incubate cu mAb la mAb indicat: S1 Braporturi molare timp de 1 oră pe gheață și supuse citometriei în flux. Suspensiile monocelulare în tampon FACS au fost centrifugate la 400 × g timp de 10 minute. Ulterior, celulele au fost incubate cu amestec S1 B și mAb timp de 1 oră pe gheață, urmate de incubare cu anticorpi anti-IgG conjugati Alexa Fluor 594 conjugați de capră anti-umană IgG (Invitrogen, Thermo Fisher Scientific, Catalog # A-11014) timp de 45 min la temperatura camerei. Celulele au fost supuse unei analize citometrice a fluxului cu ajutorul unui citometru de flux CytoFLEX (Beckman Coulter). Rezultatele au fost analizate de către FlowJo (versiunea 10). Porțile FSC / SSC au fost utilizate pentru a selecta celulele mononucleare. Colorarea anticorpilor de control a fost utilizată pentru a defini populațiile de celule pozitive / negative.
Test de neutralizare a virusului pseudotipat
Producția de VSV pseudotipată cu SARS-S și SARS2-S a fost efectuată așa cum s-a descris anterior cu unele adaptări 11 . Pe scurt, celulele HEK-293T au fost transfectate cu vectori de expresie pCAGGS care codifică SARS-S sau SARS2-S purtând o trunchiere a cozii citoplasmatice de 28 sau 18 aa. Într-o zi după transfecție, celulele au fost infectate cu VSV-G pseudotipat VSVΔG purtător de licurici ( Photinus pyralis) gena raportoare luciferaza. Douăzeci și patru de ore mai târziu, supernatanții care conțin particule de VSV pseudotipate SARS-S / SARS2-S au fost recoltate și titrate pe celulele VeroE6 de maimuță verde africană VeroE6 (ATCC # CRL-1586). În testul de neutralizare a virusului, mAbs au fost de patru ori diluate serial de două ori concentrația finală dorită în DMEM suplimentată cu 1% ser de vițel fetal (Bodinco), 100 U / ml Penicilină și 100 pg / ml Streptomicină (Lonza, Catalog nr. 17-602E ). MAb-urile diluate au fost incubate cu un volum egal de particule pseudotipate de VSV timp de 1 oră la temperatura camerei, inoculate pe monostratele VeroE6 confluente în placă cu 96 de godeuri și incubate în continuare la 37 ° C timp de 24 de ore. Activitatea luciferazei a fost măsurată pe un luminometru cu placă Berthold Centro LB 960 utilizând D-luciferină ca substrat (Promega). Procentul de infecțiozitate a fost calculat ca raport între citirea luciferazei în prezența mAbs normalizată la citirea luciferazei în absența mAb. Concentrațiile inhibitorii jumătate maxime (IC)50 ) au fost determinate folosind regresia logistică cu 4 parametri (GraphPad Prism versiunea 8).
Test de neutralizare a virusului
Neutralizarea SARS-CoV și SARS-CoV-2 autentică a fost efectuată folosind un test de neutralizare a reducerii plăcii, așa cum s-a descris anterior, cu unele modificări 23 . Pe scurt, mAbs au fost dublate serial diluate în mediu de cultură începând de la 40 ug / ml și 50 µl s-au amestecat cu 50 µl (500 TCID 50) SARS-CoV sau SARS-CoV-2 timp de 1 oră. Amestecul a fost apoi adăugat la celulele VeroE6 și incubat timp de 1 oră, după care celulele au fost spălate și apoi incubate în mediu timp de 8 ore. Celulele au fost apoi fixate și colorate folosind un ser anti-SARS-CoV de iepure (Sino Biologic) și o IgG anti-iepure marcată cu peroxidază secundară (Dako). Semnalul a fost dezvoltat folosind un substrat care formează precipitat TMB (True Blue, KPL) și numărul de celule infectate pe godeu a fost contorizat folosind analizorul de imagine ImmunoSpot (CTL Europe GmbH). Concentrațiile inhibitoare jumătate maximale (IC 50 ) au fost determinate utilizând 4 parametri de regresie logistică (GraphPad Prism versiunea 8).
Analiza ELISA a anticorpului care se leagă la antigenele spike CoV
NUNC Maxisorp placi (Thermo Scientific) au fost acoperite cu cantități echivalente de antigen la 4 ° C peste noapte. Plăcile au fost spălate de trei ori cu PBS conținând 0,05% Tween-20 și blocate cu 3% albumine bovine serice (Bio-Connect) în PBS conținând 0,1% Tween-20 la temperatura camerei timp de 2 ore. S-au adăugat patru ori diluții seriale de mAbs începând de la 10 pg / ml (diluate în tampon de blocare) și plăcile au fost incubate timp de 1 oră la temperatura camerei. Plăcile au fost spălate de trei ori și incubate cu anticorp secundar anti-uman de capră conjugat (HRP) cu peroxidază de hrean (ITK Southern Biotech) diluat 1: 2000 în blocarea tamponului timp de 1 oră la temperatura camerei. Un anticorp anti-StrepMAb conjugat HRP (IBA, Catalog nr. 2-1509-001) a fost utilizat pentru a corobora acoperirea echimolară a antigenilor cu vară marcată cu strep. Activitatea HRP a fost măsurată la 450 nanometri folosind substratul de tetrametilbenzidină (BioFX) și un cititor de plăci ELISA (EL-808, Biotek). Concentrația efectivă pe jumătate maximă (CE)50 ) valorile de legare au fost calculate prin analiza de regresie neliniară pe curbele de legare folosind GraphPad Prism (versiunea 8).
Date care stau la baza Fig. 1 b, c, 2a , Fig suplimentare. 1 , 2 , 4 și 5 sunt furnizate ca fișiere Date sursă. Secvențe de anticorpi și anticorpi sunt disponibile (contactând Vincent Rijsman de la Biroul de suport pentru cercetare al Universității Utrecht; VMCRijsman@uu.nl) în scopuri de cercetare numai în baza unui MTA, care permite utilizarea secvențelor de anticorpi în scopuri non-comerciale, dar nu și dezvăluirea acestora către terți. Toate celelalte date sunt disponibile de la autorul corespunzător la cereri rezonabile.
Referințe
1.
Zhou, P. și colab. Un focar de pneumonie asociat cu un nou coronavirus de origine probabilă a liliecilor. Natura 579 , 1–4 (2020).
Grupul de studiu Coronaviridae al Comitetului internațional pentru taxonomia virusurilor. Specia coronavirusului grav legat de sindromul respirator acut: clasificarea 2019-nCoV și denumirea sa SARS-CoV-2. Nat. Microbiol . 5 , 536–544 (2020).
4.
Prabakaran, P. și colab. Anticorpi monoclonali umani potențiali împotriva virusurilor SARS CoV, Nipah și Hendra. Opinia expertului. Biol. Ther 9 , 355-368 (2009).
Reguera, J. și colab. Bazele structurale ale atașării coronavirusului la aminopeptidaza N gazdă și inhibarea acestuia prin neutralizarea anticorpilor. PLoS Pathog. 8 , e1002859 (2012).
Prabakaran, P. și colab. Structura domeniului de legare a receptorilor coronavirusului sindromului respirator sever complexat cu anticorpul neutralizant. J. Biol. Chem. 281 , 15829–15836 (2006).
Hwang, WC și colab. Baza structurală de neutralizare de către un sindrom respirator acut anti-sever acut anticorp proteic spike, 80R. J. Biol. Chem. 281 , 34610–34616 (2006).
Rockx, B. și colab. Baza structurală pentru protecția anticorpului monoclonal uman puternic neutralizant încrucișat împotriva sindromului respirator acut letal uman și zoonotice grave provocate de coronavirus. J. Virol. 82 , 3220–3235 (2008).
Widjaja, I. și colab. Spre o soluție pentru MERS: anticorpi monoclonali umani de protecție care vizează diferite domenii și funcții ale glicoproteinei spike-coronavirus MERS. Emerg. Microbi Infect. 8 , 516–530 (2019).
Yuan, Y. și colab. Structurile Cryo-EM ale glicoproteinelor spike MERS-CoV și SARS-CoV dezvăluie domeniile dinamice de legare a receptorilor. Nat. Commun. 8 , 15092 (2017).
Gui, M. și colab. Structurile de microscopie crio-electronică ale glicoproteinei spice SARS-CoV dezvăluie o stare de conformație prealabilă pentru legarea receptorilor. Rez. Celulară 27 , 119–129 (2017).
Tian, X. și colab. Legarea potențială a noii proteine spike de coronavirus 2019 de către un anticorp monoclonal uman specific SARC coronavirus. Emerg. Microbi Infect. 9 , 382–385 (2020).
Menachery, VD și colab. Un grup de tipuri de coronavirusuri ale liliecilor în formă de SARS arată potențial de apariție umană. Nat. Med. 21 , 1508 (2015).
ter Meulen, J. și colab. Combinație de anticorpi monoclonali umani împotriva coronavirusului SARS: sinergie și acoperirea mutanților de evacuare. PLoS Med. 3 , e237 (2006).
Bosch, BJ, Bartelink, W. și Rottier, PJ Cathepsina L clivează funcțional proteina severă acută severă de coronavirus clasa I proteină de fuziune în amonte mai degrabă decât adiacentă peptidei de fuziune. J. Virol. 82 , 8887–8890 (2008).
Okba, NMA și colab. Detectarea sensibilă și specifică a răspunsurilor la anticorpi la nivel scăzut în infecțiile cu coronavirus ale sindromului respirator din Orientul Mijlociu. Emerg. Infecta. Dis. 25 , 1868–1877 (2019).
Li, F., Li, W., Farzan, M. și Harrison, SC Structura domeniului de legare a receptorului spona coronavirus SARS complexat cu receptor. Știința 309 , 1864-1868 (2005).
Mulțumim Dr. Yoshiharu Matsuura (Universitatea Osaka, Japonia) pentru furnizarea de virus VSV-G-pseudotipat cu codificare a luciferazei, și Yongle Yang, Michael van der Reijden și Rick Janssens pentru asistență tehnică. Mulțumim lui Christian Drosten (Charité Universitätsmedizin Berlin, Germania) pentru furnizarea virusului SARS-CoV-2. Acest studiu a fost realizat în cadrul Centrului Național pentru o Sănătate (NCOH), al Centrului de Imunologie Moleculară din Utrecht - Universitatea Utrecht și a Inițiativei Zoootice pentru Anticipare și Pregătire [proiectul ZAPI; acordul de grant nr. 115760]. Soareci utilizat in acest studiu au fost generate de Harbor Anticorpii BV, o companie fiica Harbor Biomed ( http://www.harbourbiomed.com). C. Wang a fost susținut de o subvenție a Consiliului Burselor Chineze (numărul de dosar CSC201708620178).
Informatia autorului
Note de autor
Acești autori au contribuit în egală măsură: Chunyan Wang, Wentao Li, Dubravka Drabek.
Acești autori au supravegheat în comun lucrarea: Frank Grosveld, Berend-Jan Bosch.
Afilieri
Secția de virologie, secția Boli Infecțioase și Imunologie, Departamentul de Științe ale Sănătății Biomoleculare, Facultatea de Medicină Veterinară, Universitatea Utrecht, Utrecht, Olanda
Chunyan Wang
, Wentao Li
, Frank JM van Kuppeveld
& Berend-Jan Bosch
Departamentul de Biologie Celulară, Centrul Medical Erasmus, Rotterdam, Olanda
Dubravka Drabek
, Rien van Haperen
& Frank Grosveld
Harbour BioMed, Rotterdam, Olanda
Dubravka Drabek
, Rien van Haperen
& Frank Grosveld
Departamentul de Virosciență, Centrul Medical Erasmus, Rotterdam, Olanda
Nisreen MA Okba
& Bart L. Haagmans
Universitatea de Medicină Veterinară, Hanovra, Germania
Albert DME Osterhaus
contribuţii
BJB a conceput, proiectat și coordonat studiul. Experiențele CW, WL, NMAO, RvH și DD au efectuat experimentele. DD, BLH și BJB au supravegheat o parte din experimente. Toți autorii au contribuit la interpretările și concluziile prezentate. BJB a scris manuscrisul, BLH, FJMK, ADMEO și FG au participat la editarea manuscrisului.
La 12 martie 2020 a fost depusă o cerere de brevet privind anticorpi monoclonali vizați de SARS-CoV-2 (cererea de brevet din Regatul Unit nr. 2003632 .3; solicitanți de brevet: Universitatea Utrecht, Centrul Medical Erasmus și Harbour BioMed). FG, DD și RvH sunt acționari non-substanțiali de interes în Harbor Biomed și au făcut parte din echipa care a generat șoarecii.
Informatii suplimentare
Informații despre evaluarea colegilor Nature Communications mulțumește recenzorii anonimi pentru contribuția lor la revizuirea inter pares a acestei lucrări.
Nota editorului Springer Nature rămâne neutru în ceea ce privește revendicările jurisdicționale din hărțile publicate și filialele instituționale.
Acces deschis Acest articol este licențiat în conformitate cu o licență internațională Creative Commons Attribution 4.0, care permite utilizarea, distribuirea, adaptarea, distribuirea și reproducerea în orice suport sau format, atât timp cât acordați credit adecvat autorului (autorilor) inițiali și sursei, furnizați un link către licența Creative Commons și indicați dacă au fost efectuate modificări. Imaginile sau alte materiale terțe din acest articol sunt incluse în licența Creative Commons a articolului, cu excepția cazului în care se indică altfel într-o linie de credit a materialului. Dacă materialul nu este inclus în licența Creative Commons a articolului și utilizarea intenționată a dvs. nu este permisă prin regulamentul legal sau depășește utilizarea permisă, va trebui să obțineți permisiunea direct de la titularul dreptului de autor. Pentru a vizualiza o copie a acestei licențe, vizitațihttp://creativecommons.org/licenses/by/4.0/ .
Wang, C., Li, W., Drabek, D. și colab. Un anticorp uman monoclonal care blochează infecția cu SARS-CoV-2. Nat Commun 11, 2251 (2020). https://doi.org/10.1038/s41467-020-16256-y